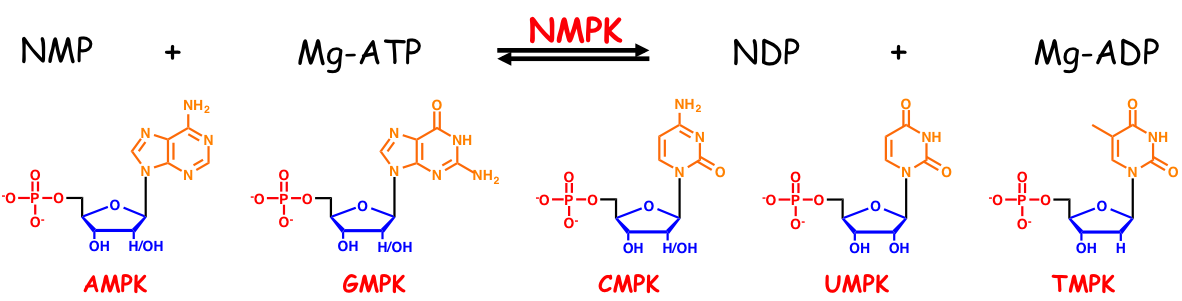
NMPKs are ubiquitous enzymes, which catalyze the reversible transfer of phosphoryl group from ATP to an NMP, and are essential for cell division, growth and metabolism. In bacteria there are five NMPKs, each one specific of a base (see scheme below), whereas, in eukaryotes, CMP and UMP are phosphorylated by a unique bi-functional enzyme, namely the UMP/CMPK.
Our research projects have been focused on NMPKs of bacterial pathogens, mainly on M. tuberculosis responsible for tuberculosis (collaboration with S. Cole and G. Marchal, Institut Pasteur) and on their human counterparts (collaboration with D. Deville-Bonne, Paris VI).
TMPK from M. tuberculosis was the first NMPK to be extensively studied. It was also used to develop and validate bioinformatics programs (coll. with D. Douguet and G. Labesse). Based on the 3D-structure of TMPK in complex with one of its natural substrate dTMP obtained in 2001 (Li de la Sierra et al, 2001), the drug design and synthesis steps (coll. with S. Pochet, S. Van Calenbergh and M. J. Perez-Perez) have been launched as well as the in silico screening of chemical libraries (coll. with G. labesse and M. Nilges). Different classes of molecules have been evaluated (see “Inhibitors of TMPK from Mycobacterium tuberculosis” project). The most promising inhibitors exhibit inhibitory potencies not only on TMPK but also on M. tuberculosis culture with MIC50 in the micromolar range. Compound optimization is still underway (coll. with S. Van Calenbergh).
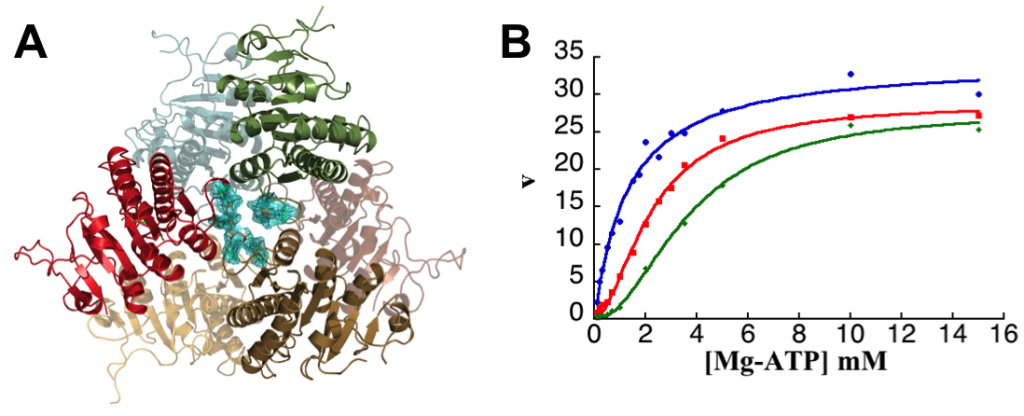
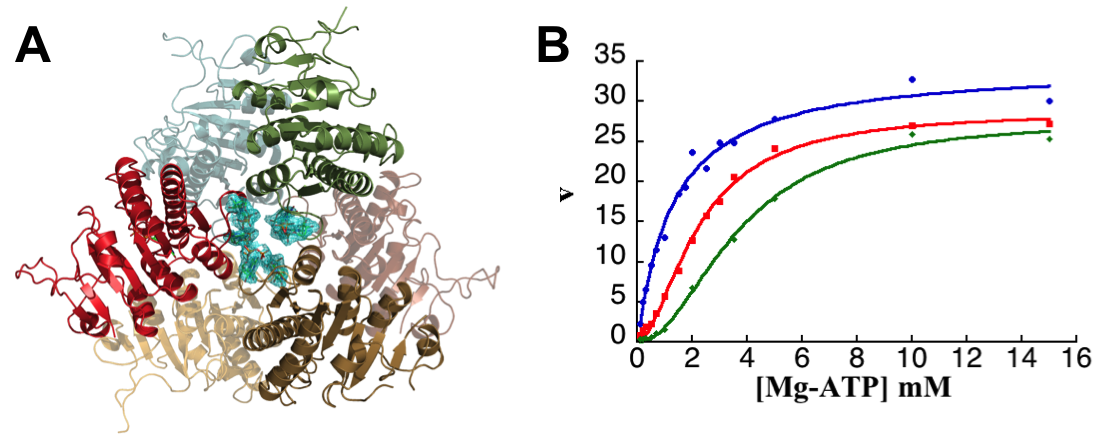
M. tuberculosis UMPK, that have been characterized more recently combining biochemical and structural investigations (Labesse et al, 2011), is another promising target as no counterpart exists in eukaryotes. The allosteric regulation of this enzyme was shown to be typical from Gram-positive bacterial UMPKs: it exhibits cooperativity for ATP and an allosteric regulation by UTP (inhibitor) and GTP (activator). To find appropriate ligands for this enzyme, chemical libraries were screened within the CBC platform (see on the “biochemistry and screening” group page) using a coupled spectrophotometric assay. Several compounds were identified as potent ligands of M. tuberculosis UMPK, one of them being a GTP antagonist or a UTP agonist, and the first example of a non nucleoside compound able to bind to the effector site(s). In the meantime, the 3D-structure of M. tuberculosis UMPK complexed with GTP was solved (in collaboration with G. Labesse) and has enabled to define the positive effector binding site. Key residues involved in the «cross-talk» between the active site and the effector binding pocket were confirmed by site-directed mutagenesis and biochemical studies. The role of macromolecular dynamics rather than major structural changes in the allosteric regulation of bacterial UMPKs was pointed out. Up to now no structure in the presence of the second effector (UTP) is available. The allosteric inhibitor identified thanks to our screening campaign will be a valuable tool to get the structure of this site that is still totally unknown for the Gram-positive bacterial UMPKs.
A. M. tuberculosis UMPK 3D-structure: bacterial UMPKs do not belong to the P-loop containing kinases such as the other NMPKs, but to the AA-kinases superfamily. B.M. tuberculosis UMPK kinetic properties showing the cooperativity toward ATP and the allosteric regulation by GTP (positive effector) and UTP (negative effector): enzyme activity was determined in the absence (red curve) or in the presence of effectors (blue curve: 0.5 mM GTP and green curve: 0.1 mM UTP).