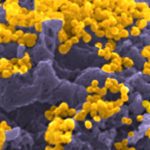



Présentation
Human Antibody Responses to Viruses
Adaptive immune responses naturally occurring upon infection or induced by vaccination generate high affinity antibodies, and memory B-cell subsets to respond to further antigenic challenges, providing protection against (re-)infections. Our research aimed at understanding the human memory B-cell responses triggered by viral infections participating to viral clearance and infection control.
The studies below were supported by the European Research Council (Grant: ERC-2013-StG HumAntiViruses).