

About
Leptospira spp. are thin, highly motile, slow-growing spirochetes that can be distinguished from other bacteria on the basis of their unique helical shape and the presence of periplasmic flagella.
We showed that peptidoglycan sacculi from Leptospira spp. retain the helical shape of intact cells. Interestingly, the distribution of muropeptides was different from that in the E. coli model, indicating that specific enzymes might be active on the peptidoglycan macromolecule. We could alter the shape of L. biflexa with the broad-spectrum β-lactam antibiotics that preferentially target penicillin-binding proteins. Loss of the MreB function was correlated with morphological abnormalities such as a localized increased diameter and heterogeneous length. These findings indicate that important aspects of leptospiral cell morphology are determined by the cytoskeleton and the murein layer, thus providing a starting point for a better understanding of the morphogenesis in these atypical bacteria .

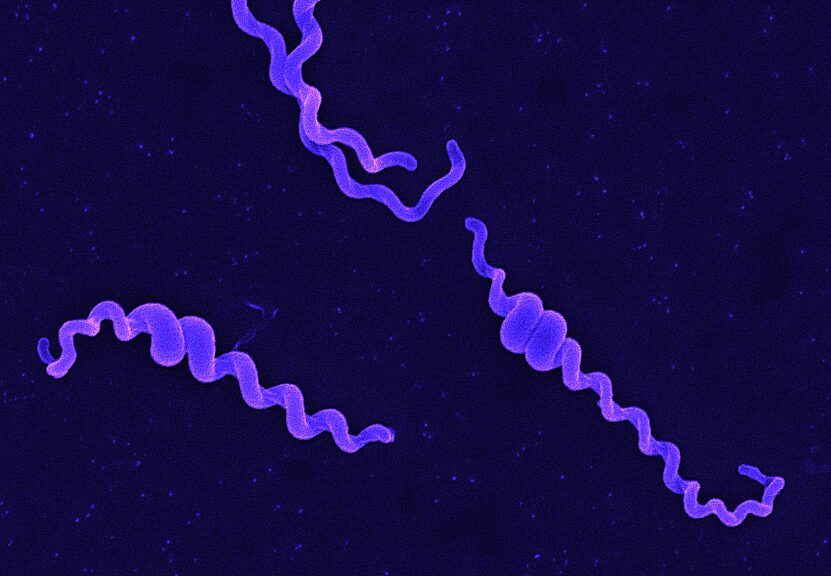
Fig. Movement of leptospires (A, from Goldstein and Charon, 1988) and electronic microscopy of Leptospira biflexa (B). During translational motility, counterclockwise rotation of the perisplasmic flagellum produces spiral-shaped ends at the leading edge while concomitantly, clockwise PF rotation causes gyrating cell ends at the tail to remain hooked-shaped
The basal body and hook structures of spirochete flagella are similar to those of the model systems of Escherichia coli and Salmonella enterica. However, in contrast to the flagellin polymer of enterobacteria, the flagellar filaments of spirochetes investigated to date have a much more complex structure.
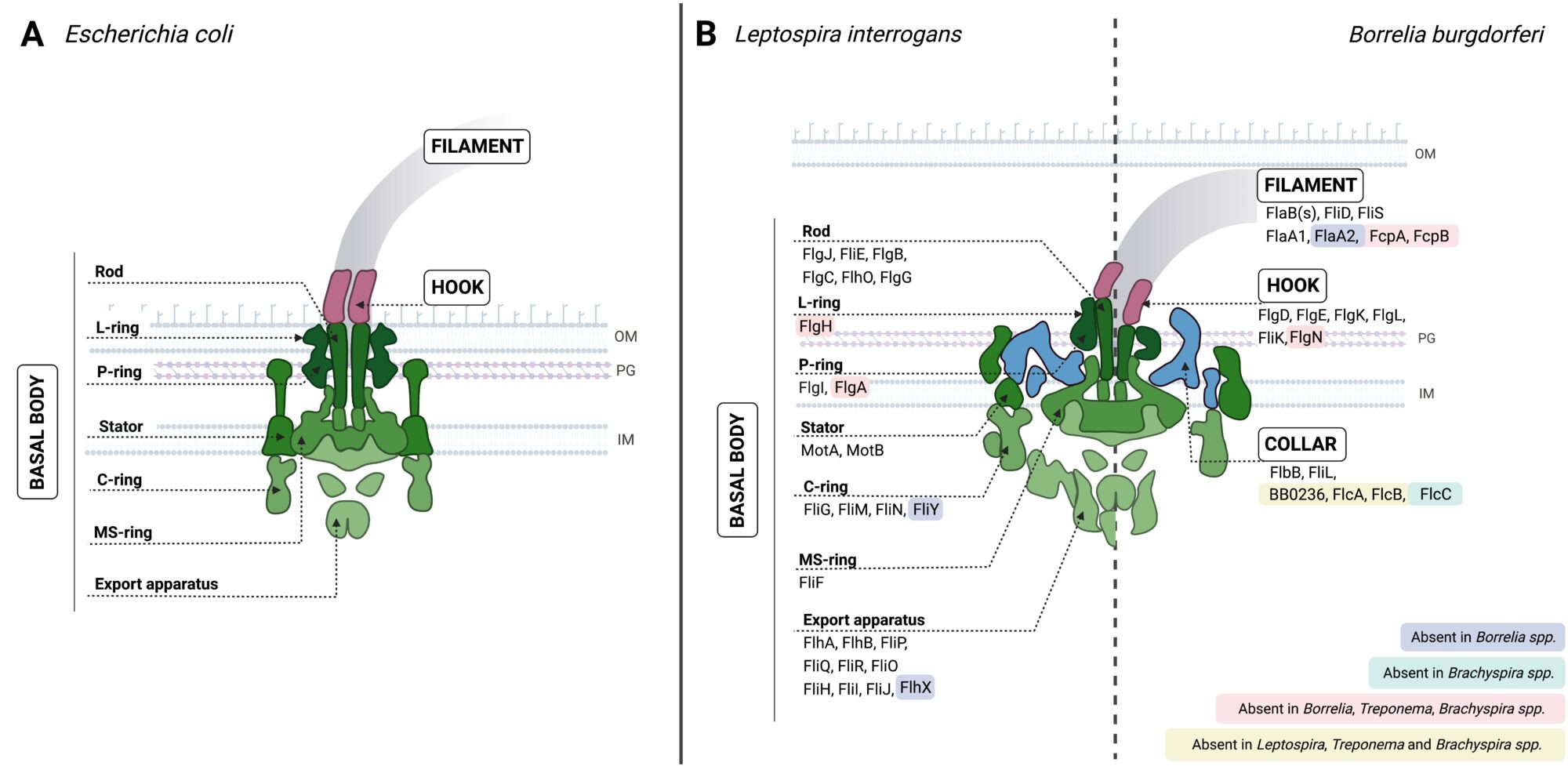
Fig. Schematic representation of the endoflagella of Borrelia and Leptospira spp. (A) and comparison with the E. coli flagellum (B).
In Leptospira, we have identified novel flagellar proteins and we have shown that the filament display an asymmetric assembly. We hypothesize that the filament’s inherent asymmetry and curvature play a key role in motility by promoting functional contacts between specific flagellar subcomponents and the peptidoglycan layer to form a loadbearing.
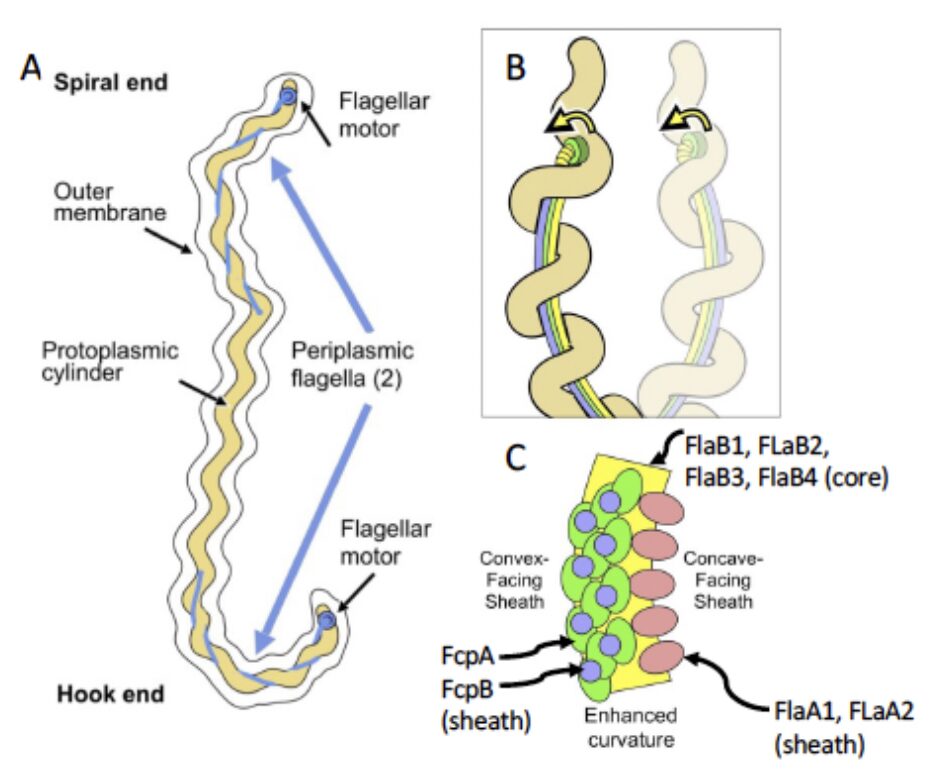
Fig. A. Rotation of the rigid and supercoiled flagellar filament in the periplasm, linked to the rolling of the cell body, is thought to propel the cell by a forward thrust and causes the characteristic hook- or spiral- shaped ends of leptospires. B. Model depicting how sheath-enforced flagellar curvature would interact with the coiled body in Leptospira to generate large-scale curved deformations in the body. C. Predicted endoflagellar filament composition shows that the filament displays an inner core composed of the flagellin homolog FlaBs surrounded by an asymmetric assembly of sheath proteins FcpA and FcpB along the convex side of the core, while FlaA1/2 bind on the concave side. Adapted from Gibson et al. 2020